脊髓性肌萎缩症(SMA),一类由脊髓前角运动神经元变性导致肌无力、肌萎缩的疾病,SMN基因与SMA密切相关。人类有SMN1和SMN2两个同源基因,SMN2虽然与SMN1高度同源,但由于其在第7个外显子发生碱基突变(C变成T),导致外显子7不包含在成熟的蛋白中,不能发挥SMN1同样的功能。目前,可通过药物Nusinersen,恢复SMN2外显子7的剪接,或者通过腺相关病毒载体递送SMN1基因的基因治疗方法,改善了小鼠模型和患者的SMA表型。尽管两种疗法均能提供显着的临床改善,但没有一种疗法可以完全治愈SMA,而且并非所有患者对治疗的反应均相同。因此,有必要对SMN表达调控机制进行更深入的探索。
越来越多的研究表明,circRNAs在基因调控中具有重要的作用。促进circRNAs生物发生的主要机制是反向重复序列的存在,尤其是反向的Alu重复序列。此外,识别内含子区域的RNA结合蛋白(RBP)的二聚化也可以促进circRNA的生物发生。然而,目前尚不清楚RBP是否利用Alus来介导circRNAs的生物发生。值得一提的是,SMN1基因是人类基因组中Alus元件丰度最高的基因之一,结合Alus在circRNA生成作用中的机制认识,SMN1基因来源的circRNA生成过程与这个基因的表达和功能有什么关系呢?
2019年11月28日,意大利天主教圣心大学Claudio Sette教授实验室在Nucleic Acids Res(中科院JCR一区,IF=11.147)杂志上发表了一篇题为Sam68 binds Alu-rich introns in SMN and promotes pre-mRNA circularization 的研究论文,揭示了RBP蛋白Sam68通过结合SMN mRNA前体中的Alus,促进circRNA的生物发生的分子机制。

该研究首先发现SMN基因中存在大量的Alus元件,其中有很多以反向排列,形成反向互补序列。进一步,作者通过沉默细胞中RNA解旋酶DHX9,增加SMN mRNA前体反向互补配对的几率,发现这些反向互补序列的配对负调控SMN基因的转录进程。
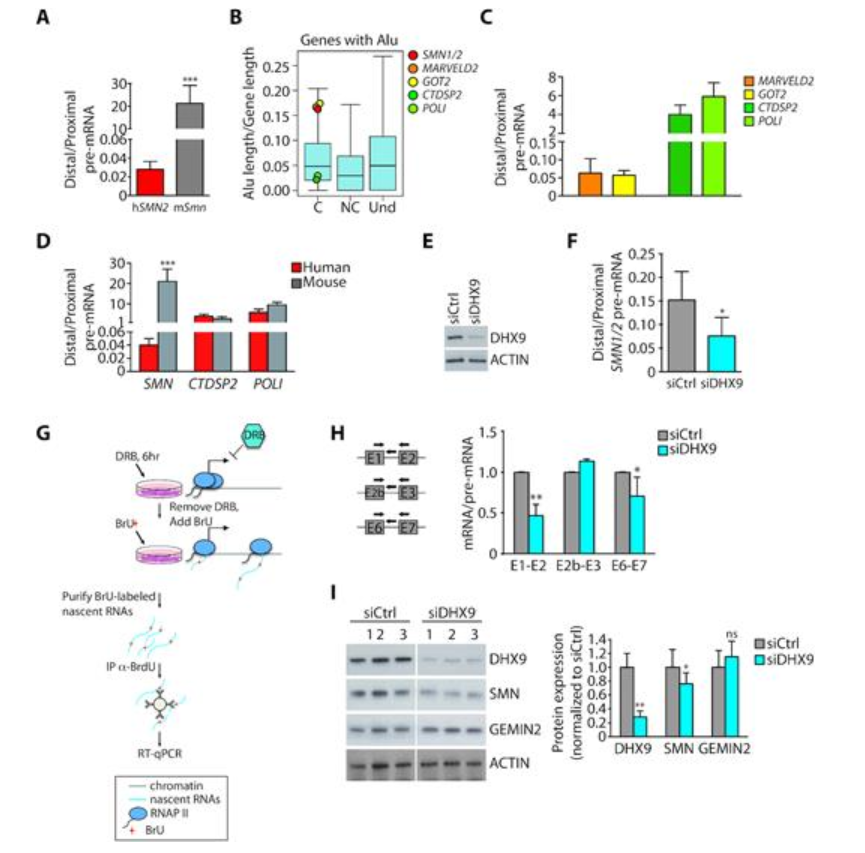
图1 反向重复Alus元件负调控SMN基因转录
接下来,研究发现,干扰DHX9后,起源于SMN基因的circRNAs生物发生减少,SMN蛋白丰度增加,所以推测减少SMN 来源circRNAs的生成是否可以恢复SMN蛋白的含量。为了检测这一假设,该研究使用ASO(an antisense oligonucleotide),干扰了由SMN外显子8内隐性剪切位点形成的circRNA的形成,结果发现在细胞中转染ASO后,SMN前体mRNA的转录没有发生变化,但SMN蛋白的丰度明显地增加了20%,表明cicrRNA与基因座的蛋白质编码进行潜在的竞争。
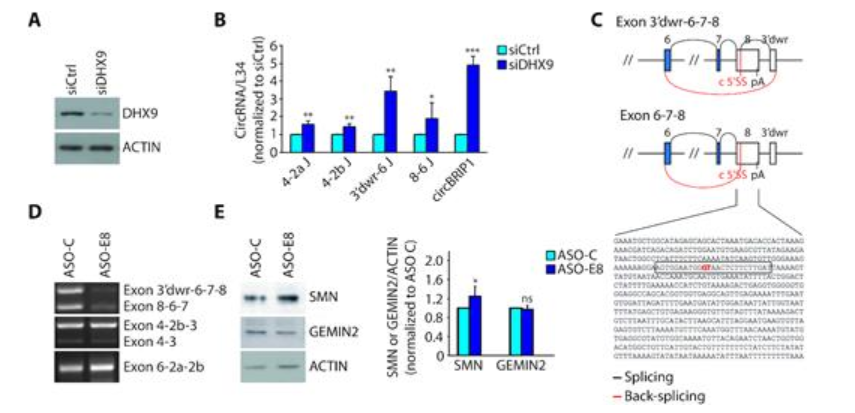
图2 干扰SMN circRNA的生物发生影响SMN蛋白的表达
考虑到RBP蛋白可以识别内含子区域,从而促进circRNAs的生物发生,该研究在这些序列中搜索了RBP特异性基序,并确定了四个RBP的强结合位点:Sam68,TIA-1和hnRNP F / H。敲除实验表明,只有敲除Sam68导致几种SMN circRNA的表达显著降低。随后通过CLIP(UV crosslink immunoprecipitation)实验进一步表明Sam68和SMN基因的内含子强烈结合。
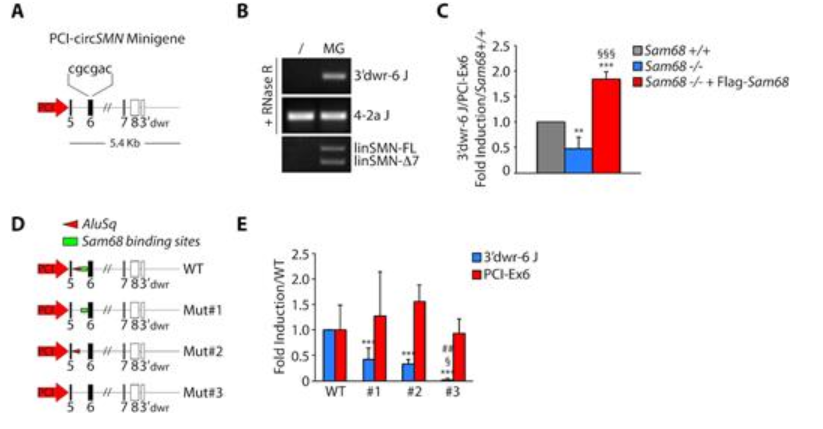
图3 Sam68与富含IRAlu的内含子互作
Sam68将单链富含A / U的二聚体基序识别为目标转录本中的二聚体。由于这种高亲和力结合位点在Alus侧翼区域也非常丰富,因此该研究假设Sam68与这些Alu近端区域的结合及其同源二聚化可能有助于使位于遥远内含子中的IRAlus更加接近,从而有利于circRNA生物发生。通过缺失突变反向互补序列或RBP结合位点,发现两者均会影响circRNAs的生物发生,并且同时缺失Alu和Sam68结合区会引起累加效应,表明Sam68与IRAlu配对合作促进SMN 前体mRNA的环化。
综上所述,该研究发现SMN内含子中富含丰富的Alu元件,并可以形成反向互补配对,促进SMN 的环化。另外,该研究还发现RBP蛋白Sam68可以与SMN 前体mRNA中的内含子互作,也促进circRNA生物发生。总之,该研究揭示了SMN基因座的新型调节层,可能对SMA产生影响。
参考文献
Pagliarini, V., Jolly, A., Bielli, P., Di Rosa, V., De la Grange, P., & Sette, C. (2019). Sam68 binds Alu-rich introns in SMN and promotes pre-mRNA circularization. Nucleic acids research.