8月28日,Nucleic Acids Research(IF=11.147,中科院JCR 1区)在线发表了Thomas J. Sanford团队的一篇关于circRNAs研究论文,报道了黄病毒基因组RNA的环化抑制了从头翻译起始过程。

简介
黄病毒,包括登革热病毒(DENV)和黄热病病毒,在人类中引起严重疾病,而母亲感染寨卡病毒(ZIKV)后会诱发新生儿小头畸形。感染病毒后,黄病毒RNA基因组既要被翻译产生蛋白,也必须作为新基因组转录的模板。翻译所需的核糖体和转录所需的病毒聚合酶沿基因组RNA以相反的反向起始,存在碰撞和复制失败的风险。虽然黄病毒基因组RNA通常是线性的,但其5’和3’UTR区可以碱基配对,促使基因组RNA形成环状,而5’和3’UTR区碱基配对对于其复制是必需的。本实验使用体外重建的方法,证明黄病毒ZIKV和DENV基因组RNA的环化抑制其从头翻译起始,而线性构象是翻译能力的。本实验的结果表明,清除病毒的核糖体可以促进有效地复制,并验证了黄病毒基因组的线性和环状构象的相反作用。
ZIKV病毒翻译起始的体外重建
为了研究黄病毒翻译的机制,作者在体外重建了ZIKV 基因组RNA的翻译起始体系。这种方法以前曾用于成功探索细胞戴帽结构依赖性和病毒IRES(核糖体进入位点)依赖性翻译的机制。通过兔网织红细胞裂解物或大肠杆菌获得EIF(真核翻译起始因子)和小核糖体亚基的纯化蛋白。将EIF与体外转录的RNA(携带ZIKV病毒的 5’UTR和衣壳蛋白前252个核糖核苷酸,ZIKV5’utr+)一起温育。由于组装的48S复合物可以阻止逆转录酶,导致在起始密码子下游15-17nt处产生截短的cDNA产物,因此可以通过引物延伸抑制实验以测定48S复合物的组装。该技术的示意图如图1B所示。
作者研究了5’戴帽结构对重构系统中ZIKV病毒基因组 RNA翻译起始效率的影响。 在存在完整的标准翻译起始因子的情况下,当ZIKV5’utr+ RNA未加帽时,可以检测到非常少的48S复合物形成。RNA的加帽导致在真正的AUG108处形成48S复合物。
作者接下来试图通过对加帽的ZIKV5’utr+ RNA进行系统的缺失实验来确定哪些eIF是ZIKV翻译起始所必需的。eIF2和eIF3的缺失完全抑制了AUG108的翻译起始,而在没有eIF4F的情况下会产生非常少的48S复合物,与5’戴帽结构的翻译一致。无论有没有可以在1型和2型小核糖核酸病毒核糖体进入位点翻译起始的eIF4G736-1115片段时,均不能弥补缺失eIF4F的状况。但是,使用相同的纯化蛋白,当仅包含小核糖体亚基,eIF2和Met-tRNAiMet时,就可以检测到来自猪瘟病毒HCV-like 核糖体进入位点的48S复合物组装,确认这些蛋白足以起始黄病毒科其他属病毒的核糖体进入位点的翻译。缺少eIF1时,ZIKV翻译起始同样不能进行。然而,作者观察到eIF1A的缺失轻微减少了AUG108的起始,但促进了上游的近同源密码子的选择,特别是UUG41和更小程度的UUG86。因此,虽然启动时绝对需要eIF1,但仍需要eIF1A来保持选择的准确性。
由于黄病毒5’UTR高度结构化,因此作者研究了RNA解旋酶对ZIKV翻译起始的影响。除了eIF4F复合物中存在的eIF4A之外,在加帽的ZIKV5’utr+ RNA上进行48S复合物组装需要额外的eIF4A。同样,缺失eIF4B(一种eIF4A辅助因子,可以与黄病毒基因组RNA结合)也降低了翻译起始效率。eIF4A抑制剂hippuristanol也以剂量依赖方式减少48S复合物的组装。该研究与最近的一项研究ZIKV在A549细胞和原代人肝细胞中的复制被另一种eIF4A抑制剂silvestrol抑制是一致的。
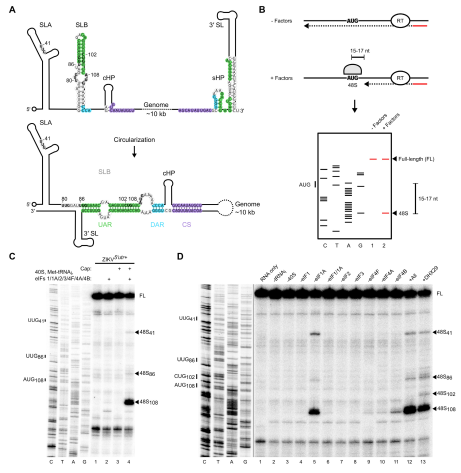
图1 ZIKV RNA上的48S复合物装配依赖于戴帽结构
ZIKV复制需要基因组环化
先前已报道DENV和WNV(西尼罗河病毒)病毒复制依赖于完整的5’和3’ UTR RNA-RNA相互作用,但这些对于翻译是不必要的。因此,作者研究了突变3’环化序列(CS)后对ZIKV病毒复制和翻译的影响。作者将报告基因Nluc插入到ZIKV基因组RNA中,并突变3’ CS序列,转入Vero细胞中,并以在病毒聚合酶活性位点突变G664DD为G664AA作为阴性对照,并通过qPCR定量基因组拷贝数,同时测量荧光素酶以监测病毒基因组的翻译。
所有RNA的基因组拷贝数在电穿孔后6和24 h之间降低,48小时后,WT RNA回升,但ZIKVNluc-GAA突变体不能复制。ZIKVNluc-Δ3’CS突变体RNA与ZIKVNluc-GAA突变体一样,转入细胞后ZIKV不能进行复制,表明ZIKV病毒复制需要完整的5′-3’环化序列的相互作用。病毒基因组的翻译与RNA复制的动力学一致。重要的是,转入细胞6 h后,此时转入WT RNA、ZIKVNluc-GAA和ZIKVNluc-Δ3’CS细胞中ZIKV的基因组拷贝数是一致的,并且这三种细胞中荧光素酶的量相似,表明WT和Δ3’CS突变体在该实验系统中RNA以相同的效率翻译。
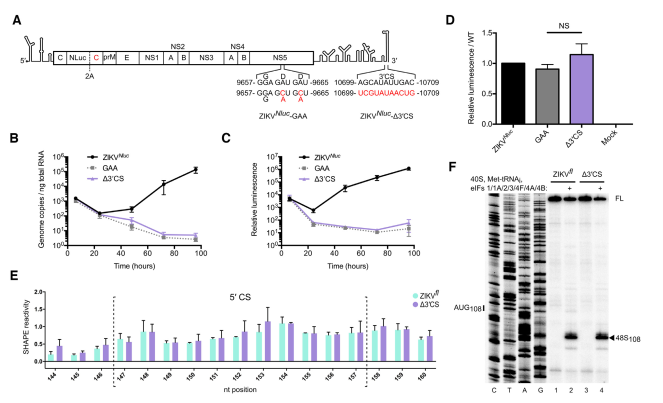
图2 ZIKV复制需要基因组环化
环化抑制ZIKV从头翻译起始
最近的一项研究表明,通过RNA结构分析,感染期间细胞中大多数ZIKV基因组RNA是线性的,对于在细胞中检测基因组环化对翻译的影响是不利的。为了克服这个问题,作者使用体外重建技术直接比较受控系统中线性和环状模板的翻译。作者首先进行体外转录,并为缺乏衣壳复制或Nluc报告基因的WT全长ZIKV基因组RNA(ZIKVfl)加帽。由于该RNA含有完整的5’和3’UTR,作者假设它们可以配对并提供合适的模板用于评估环化对翻译起始的影响。为了确定是否是这种情况,作者使用SHAPE(selective 2′-hydroxyl acylation and profiling experiment)分析RNA结构。
SHAPE分析显示体外转录的ZIKVfl RNA采用线性构象。作者假设减少UTR的~10kb可能增加环化的可能性。因此,作者构建了ZIKV小基因组(ZIKVmini),其在未改变的5’和3’个UTR中间加上60 nt的接头。以ZIKVmini为背景,构建了突变体,如图所示。首先通过天然PAGE检查ZIKVmini的构象。环状RNA的结构更紧凑,因此比线性RNA迁移更快。 一致地,ZIKVmini- 3 ’CS和ZIKVmini-5’CS比WT ZIKVmini迁移慢。而ZIKVmini-CS/Swap与WT迁移速度一样,表示,ZIKVmini- 3’ CS和ZIKVmini-5’ CS RNA为线性的,ZIKVmini-CS/SwapRNA中有碱基配对。
随后,通过SHAPE进一步分析了ZIKVmini RNA的结构,发现WT ZIKVmini的5’CS区核苷酸中具有非常低的反应性,而ZIKVmini-Δ3’CS中的5’CS区的核苷酸是高度反应性的,表明WT ZIKVmini RNA存在碱基配对,而ZIKVmini-Δ3’CS中不存在碱基配对。在ZIKVmini-CS/Swap的5’CS区内,核苷酸显示出与WT ZIKVmini类似的降低的反应性,表明交换5’和3’CS元件恢复碱基配对。结合PAGE和SHAPE两个实验结果表明,WT ZIKVmini 和ZIKVmini-CS/Swap是碱基配对的,而ZIKVmini-Δ3’CS是线性的。
作者接下来在加帽的ZIKVmini RNA上进行体外重建。与WT ZIKVfl RNA相反,在WT ZIKVmini的起始AUG108处检测到少的48S复合物组装。5’或3’CS元件的突变(RNA不能环化),增加了该位点的48S复合物的形成,同时ZIKVmini-CS/Swap(5’CS和3’CS交换,可以环化)中AUG108处也检测到较少的48S复合物组装,表明对翻译起始的影响不是由于突变,而是由于基因组环化。与CS区突变类似,5’或3’UAR序列的突变也刺激了48S复合物的形成,而5’-3’ UAR交换再次逆转了这种影响。
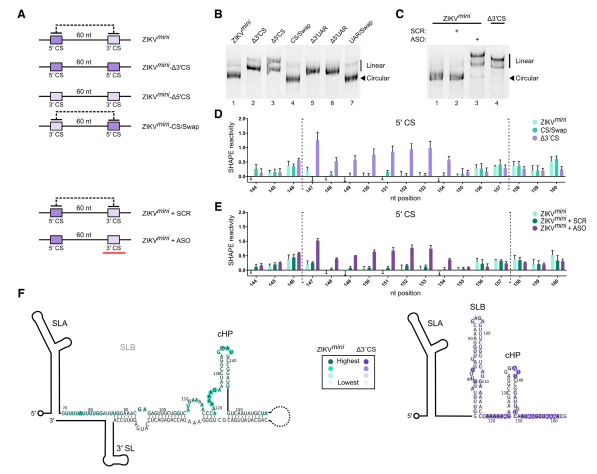
图3 ZIKVmini RNA被环化
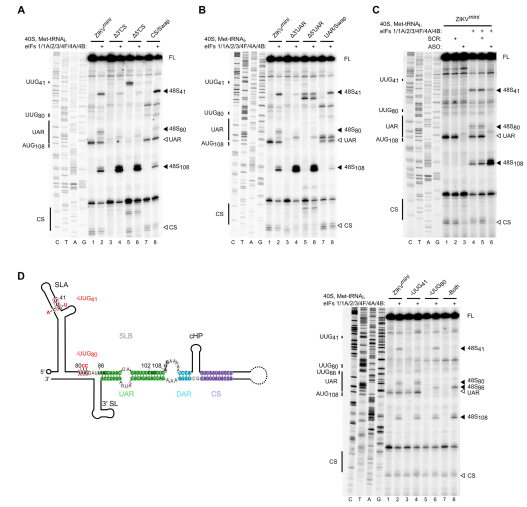
图4 ZIKV基因组环化抑制翻译起始
基因组环化损害了翻译起始的扫描过程
有趣的是,虽然ZIKV RNA环化在AUG108减少了48S复合物组装,但两个上游近同源密码子(UUG41和UUG80)的48S复合物组装得到增强,表明环化的ZIKV RNA导致翻译过程中的扫描紊乱。
作者接下来检查了在环化WT ZIKVmini的背景下突变UUG41和/或UUG80的影响。 引入的突变显示在图4D中。UUG80突变为UCC,而UUG41被UUU取代,具有补偿性C21A突变以维持SLA中的碱基配对。引入的突变消除了UUG41和UUG80处的48S复合物组装。有趣的是,UUG41和UUG80的同时突变在AUG108没有增强48S复合物组装,而是选择了UAR的5’前缘的UUG86。这些数据表明UUG41或UUG80的起始并没有严格抑制环化RNA构像中AUG108处的 48S复合物的组装,单独的RNA结构就足以介导这种现象。
基因组环化还抑制DENV中的从头翻译起始
由于环化是所有黄病毒的共同特征,所以作者研究了环化是否也影响DENV的翻译起始。作者检测了DENV1或DENV4的5’和3’UTR的mini基因组RNA的48S复合物组装。作者用与ZIKVmini RNA的方法相似的方法,通过突变3’ CS或添加与3’环化元件互补的短ASO来破坏基因组环化。与ZIKV一样,环化抑制DENV1mini和DENV4mini基因组RNA的从头翻译起始,而环化的破坏增强了真实起始AUG上的48S复合物形成(DENV1中的AUG95和DENV4中的AUG102。与ZIKV一样,观察到DENV RNA的上游UUG48的增强选择,与环化形式的扫描缺陷一致。总的来说,作者的数据表明基因组环化是不同的黄病毒用于控制基因组使用的保守机制。
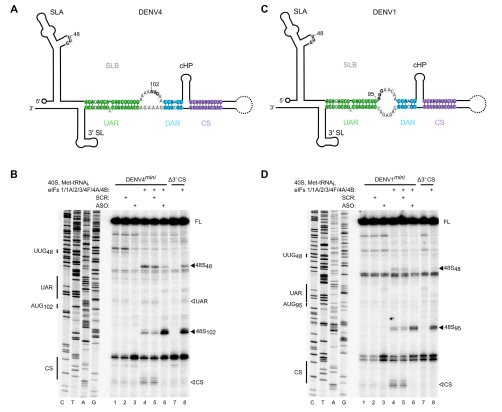
图5 环化抑制DENV基因组RNA的翻译起始
参考文献
Sanford, T. J., Mears, H. V., Fajardo, T., Locker, N., Sweeney, T. R. (2019). Circularization of flavivirus genomic RNA inhibits de novo translation initiation. Nucleic Acids Research. DOI: 10.1093/nar/gkz686