图片摘要:RNA结合蛋白UPF1和G3BP1可以选择性地介导结构复杂circRNA的降解。
2020年4月15日,四川大学华西医学院的彭勇教授和魏霞蔚教授为共同通讯作者在Trends in Cell Biology (IF=16.588)杂志上发表了一篇题为“Structure-Mediated Degradation of CircRNAs”的亮点介绍(SpotLight),介绍最近在Molecular Cell报道的由RNA结合蛋白UPF1和G3BP1选择性介导结构复杂circRNA降解的新机制([1])。circRNA公众号也曾在第一时间对该进展做了介绍:
推荐阅读:Molecular Cell:RNA降解的新机制,对circRNA也起作用

环状RNA(circRNA)是由大量真核基因产生的单链共价闭合RNA环。
它具有组织或发育阶段特异性,在生理和病理过程中都发挥着重要作用。越来越多的证据表明,circRNA通过不同的机制在转录、转录后和翻译水平上控制基因表达,包括作为miRNA海绵,与蛋白质或DNA相互作用以及编码小肽。调控circRNA的产生和降解,对于维持它的细胞内稳态和发挥它的生物学功能是至关重要的,但是circRNA的降解机制尚不清楚。本篇综述回顾了4种circRNA的降解形式,及阐述结构介导circRNA的降解途径。
miRNA以序列依赖性的方式介导AGO2对circRNA的降解
由于circRNA上没有5′-7-甲基鸟苷帽或3′-poly(A)尾巴,因此它可能是通过核酸内切酶诱导的降解。circRNA CDR1as / ciRS-7上存在与miR-671几乎完全互补匹配的位点,募集miR-671,引起AGO2核酸内切酶的切割从而破坏环状结构,再由核酸外切酶对线性RNA进行降解。然而,尚未发现是否其他miRNA介导AGO2降解带有miRNA结合位点的circRNA。
核酸内切酶RNase P / MRP以序列依赖性方式切割含m6A的circRNA
含m6A的circRNA可以使用m6A作为标记物来募集YTHDF2蛋白和HRSP12蛋白。然后,HRSP12直接与circRNA上的GGUUC模式结合,并充当将YTHDF2和核酸内切酶RNase P / MRP结合在一起的桥梁,从而使RNase P / MRP能够启动circRNA的降解。
RNase L依赖circRNA的结构调节其降解
circRNA独特的空间结构使它们能够结合各种蛋白质,从而增加了其降解机制的复杂性和特异性。许多circRNA会形成16-26个碱基对(bp)的RNA双链体,其中的碱基并不是全部配对结合,该双链体与双链RNA(dsRNA)激活蛋白激酶PKR相关。病毒感染后,RNase L(一种广泛表达的胞质核酸内切酶)通过未知机制被激活并降解这些circRNA,从而激活PKR进行先天免疫应答。然而,关于RNase L是如何被募集到circRNA中的,以及circRNA通常是如何以结构依赖的方式降解的,人们知之甚少。
UPF1和G3BP1依赖circRNA的结构调节其降解
在最近的一项研究中,Fischer等人发现了一种RNA降解机制,在正常状况下,高结构的RNA,包括circRNA被选择性的降解。人类三分之一的circRNA被预测可以形成高度整体结构,它们的降解受到两种RNA结合蛋白UPF1和G3BP1的调控。UPF1和G3BP1负责识别和解开circRNA的结构。这种降解途径需要UPF1的RNA结合活性和解旋酶活性,以及G3BP1的RNA结合活性和S149磷酸化(与其核酸内切酶活性有关)。由于这种途径感知的是整个RNA结构,而不是线性的一级结构,所以它被称为结构介导的RNA降解(SRD)。SRD不同于其他UPF1介导的RNA降解,如NMD和Staufen1介导的降解(SMD),因为这两条途径关键因子的丢失对高结构circRNA的表达几乎没有影响。
生化实验表明,G3BP1只与高结构的circRNA结合,而UPF1既可以与高结构的circRNA结合,也可以与低结构的circRNA结合。此外,在G3BP1基因敲除细胞中进一步敲除UPF1,对高结构的circRNA没有额外的影响,因此表明G3BP1是SRD途径的决定因子。
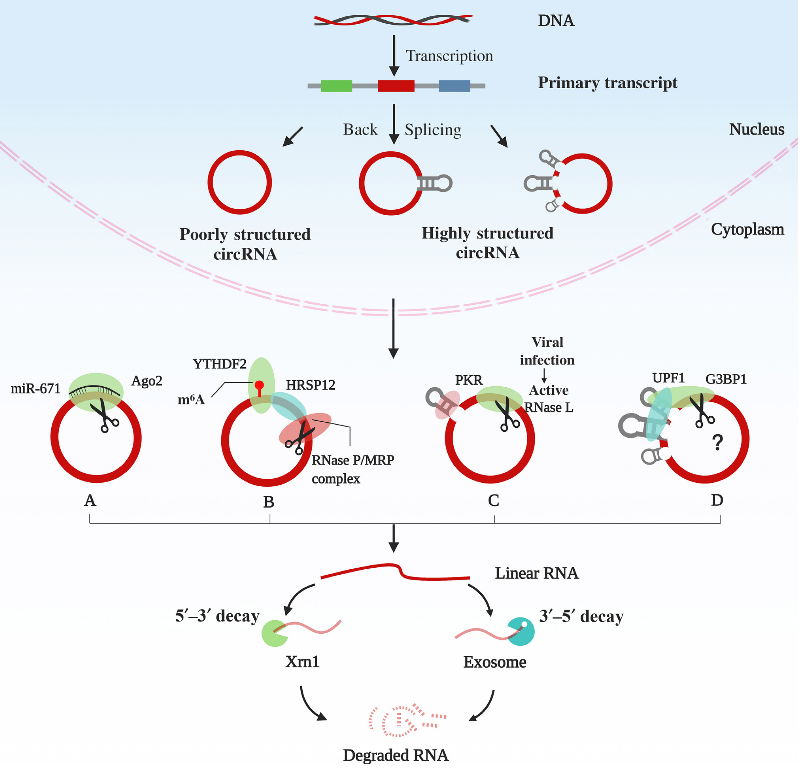
图1 circRNA的降解途径
参考文献:
[1] YingliGuo, XiaweiWei, and YongPeng. Structure-Mediated Degradation of CircRNAs. Trends Cell Biol. 2020 Apr 15. pii: S0962-8924(20)30072-6. doi: 10.1016/j.tcb.2020.04.001.