7月26日,Molecular Cell在线发表了杨力,陈玲玲教授撰写的综述文章,系统汇总了目前circRNA生成机制,功能机制以及研究面临的困难等[1]。

随着circRNA研究的不断深入,一些相关的基本科学问题得到了解决,但依然存在一些尚未解释清楚的重要问题,在这篇综述中,作者详细汇总了circRNA生成机制,功能机制的进展,针对circRNA研究所面临的困难和挑战也做了系统的总结。下面我们就一起学习一下这篇综述吧:
circRNA的生成机制
目前已报到的circRNA绝大部分来源于具有编码蛋白功能的mRNA,少数来自非编码RNA或基因间序列。circRNA生成机制的认识总结而言包括以下几个方面:
(1)circRNA存在特殊的可变剪切形式。可变剪切是mRNA功能多样性的重要机制,在circRNA中也存在可变剪切方式,四种类型的可变剪切方式都存在,有趣的是circRNA往往具有mRNA不具备的特殊剪切产物。导致这些特殊剪切产物的机制还不清楚。
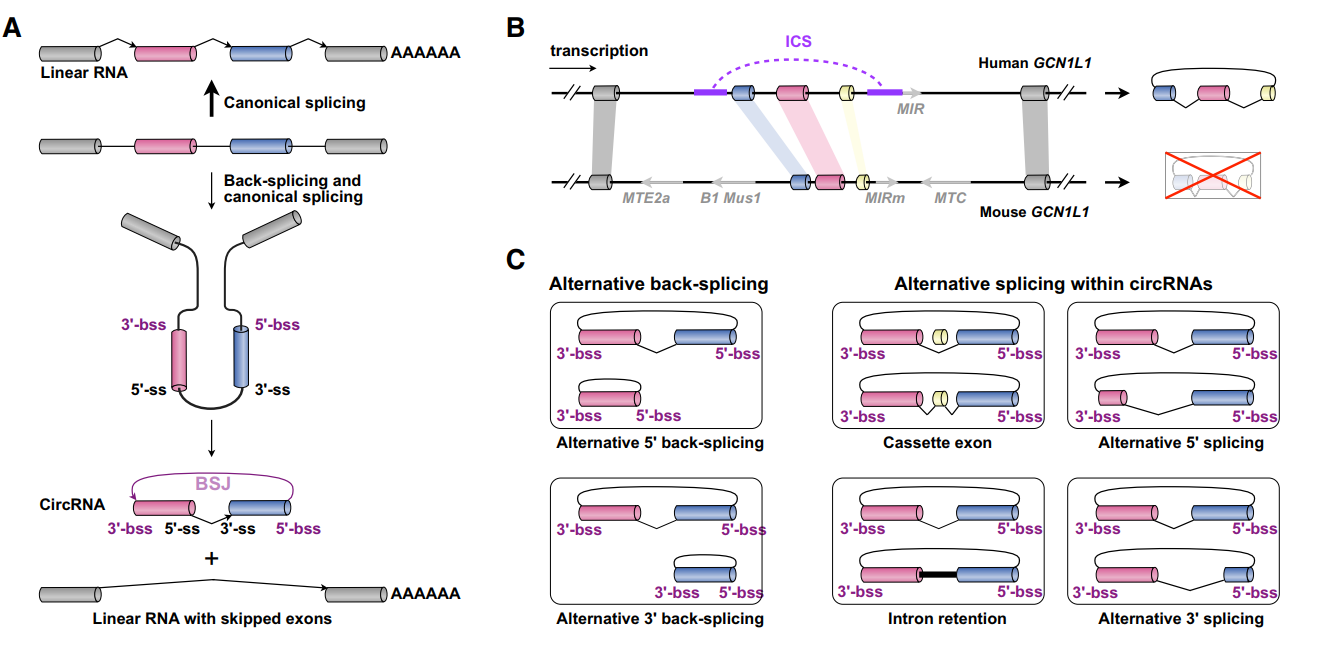
图1 circRNA存在特殊可变剪切方式 (来自[1])
(2)circRNA生成与host gene的mRNA成熟过程存在关联。circRNA与对应host gene的mRNA来源于同一转录产物,circRNA的生成可以伴随着转录过程产生,也可以在转录后加工过程中形成,后者占多数。circRNA的生成速率明显低于对应mRNA。转录延伸速度显著影响circRNA生成的效率,转录速率快有利于circRNA的形成。circRNA的生成过程依赖于剪切体,但剪切体组份异常能促进circRNA的形成。
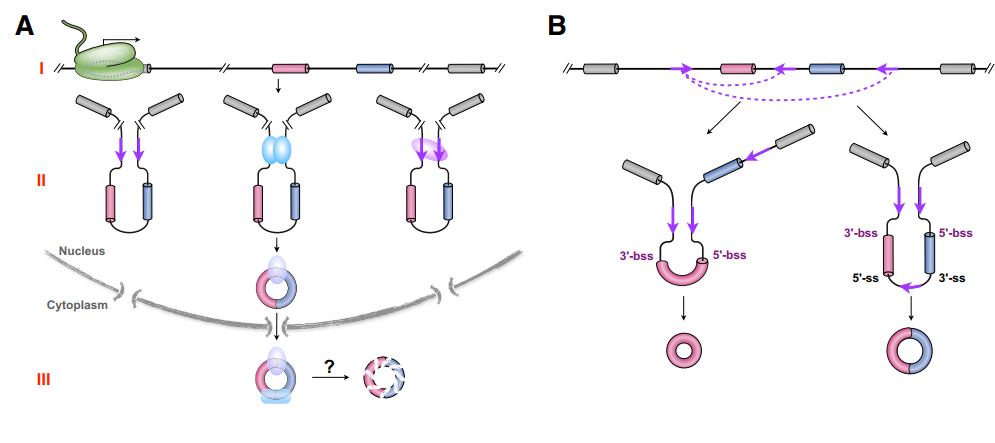
图2 circRNA生成的调控是多层次的 (来自[1])
(3)circRNA的生成可以受到顺式作用元件和反式作用因子的调控。circRNA的形成过程可以由反向拼接外显子两侧内含子中的反向互补元件驱动,如灵长类的Alu元件,小鼠中的SINE等等。也可以由能结合两侧内含子中特殊序列或结构的RNA结合蛋白驱动完成,已报到的促进circRNA生成的蛋白因子包括:双链RNA结合蛋白(如NF90/NF110,DHX9,ADAR1),也可以是结合特殊序列的蛋白(如QKI,MBL(果蝇),Fus,HNRPL,RBM20等)。
circRNA与mRNA的成熟过程都依赖于剪切体,但剪切体。
(4)circRNA的降解机制认识还十分有限。circRNA分子相对于线性的RNA分子更稳定,能一定程度耐受核酸外切酶的降解作用。circRNA分子的半衰期明显长于常规的线性RNA。但关于circRNA如何在体内降解的机制目前还知之甚少。miR-671介导的AGO2降解CDR1as是目前研究最清楚的circRNA降解机制模型,细胞内是否有类似的机制还需要进一步的探索。此外一个比较能接受的假设是细胞内的RNA内切酶介导了circRNA的降解,但目前还没有找到实例证明这一假设。作者还假设circRNA中的m6A修饰可能参与了circRNA的降解作用。
circRNA功能模型
目前circRNA的功能模型已有比较成熟的认识,概括而言,包括:竞争性结合miRNA的“miRNA Sponge”模型;竞争性结合蛋白的“protein sponge”模型;直接翻译产生蛋白;体内反转录后融合至基因组,形成假基因;定位于核内的circRNA参与调控基因的转录和RNA剪切过程。cricRNA与众多人类疾病存在相关性,相关的研究非常多。
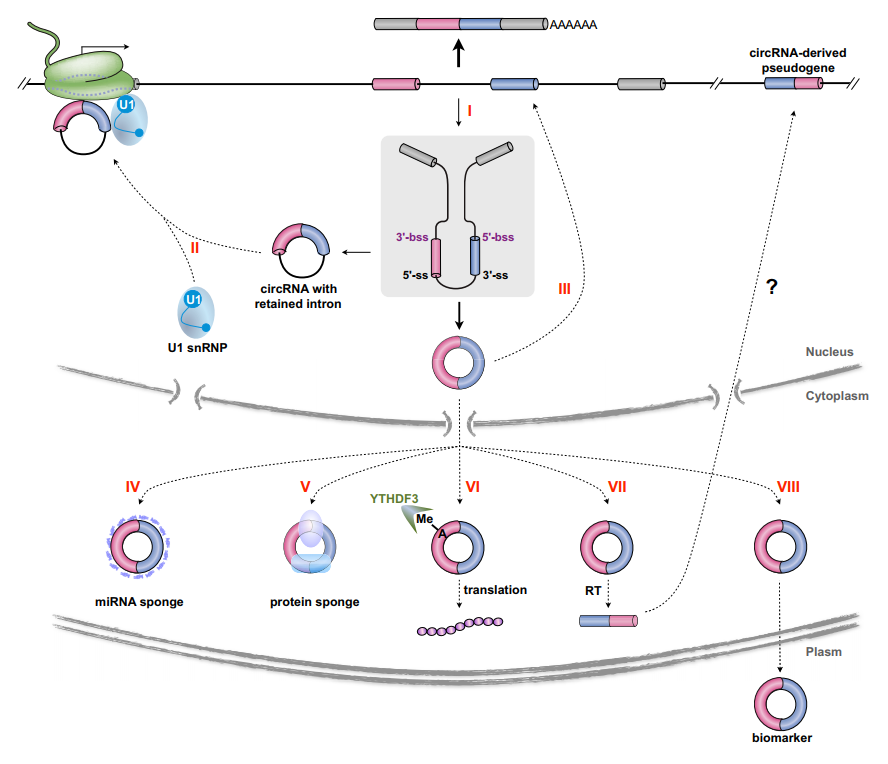
图3 circRNA功能模型(来自[1])
目前常用circRNA研究技术
目前已形成一些circRNA研究的技术体系,包括:测序与生物信息学分析circRNA表达组;跨接口的引物扩增鉴定特定circRNA分子;干扰RNA或反义核苷酸(ASO)进行Knock-down;过表达载体进行过表达;基于Cas-9打靶技术进行全circRNA序列敲除或者内含子中反向互补序列的敲除;基于Cas-13a或Cas-13b的RNA干扰技术;基于FISH的亚细胞定位检测等等。但大部分研究技术还需要进一步优化,包括RNA干扰效率,基因敲除策略,过表达载体优化等一系列问题仍需要进一步提高。
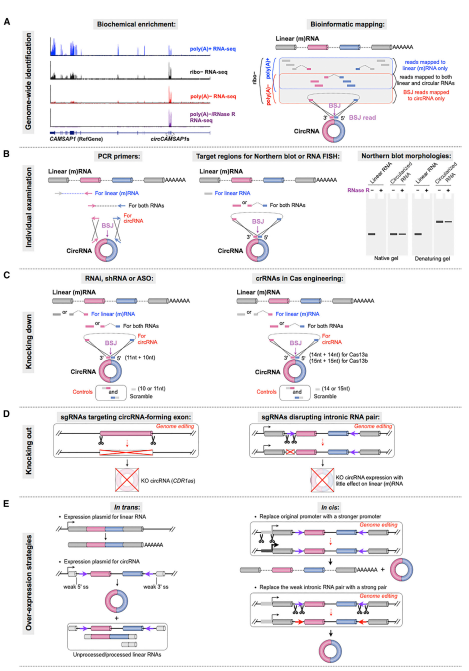
图4 circRNA研究技术(来自[1])
circRNA研究最大困难在于与对应线性mRNA的序列高度一致,仅有接口位置存在的反向拼接位点的差别,这给circRNA的研究带来了不小的麻烦和挑战。此外,内源性的circRNA普遍丰度很低,也为相关的研究带来了一些困难和挑战。不过相信随着技术体系的日益完善,相关的困难和障碍一定能得到进一步的解决。
参考文献:
1. Xiang Li, L.Y.a.L.-L.C., The Biogenesis, Functions, and Challenges of Circular RNAs. Molecular Cell, 2018.