非编码RNA(ncRNA)可以调节各种肿瘤过程,是重要的潜在癌症诊断和预后生物标志物。通常认为,它们不编码蛋白质,但生物信息学和翻译组学已经开始阐明由ncRNA编码的功能肽的作用和作用方式。近日,中南大学肿瘤研究所熊炜教授团队在Molecular Cancer(IF=10.679,中科院1区)杂志上发表了一篇题为‘Emerging role of tumor-related functional peptides encoded by lncRNA and circRNA’的文章,从预测方法和分子机制方面系统总结了肿瘤相关长链非编码RNA(lncRNA)和环状RNA(circRNA)编码短肽的最新进展。

ncRNA编码潜力的预测和小肽的鉴定
鉴于对ncRNA编码的多肽的兴趣日益增加,已经开发了许多预测和实验鉴定方法来确定ncRNA的编码能力。作者表明,这些方法包括阅读框(ORF)预测,翻译起始成分(IRES)预测,保守性分析以及翻译组学和蛋白质组学等。
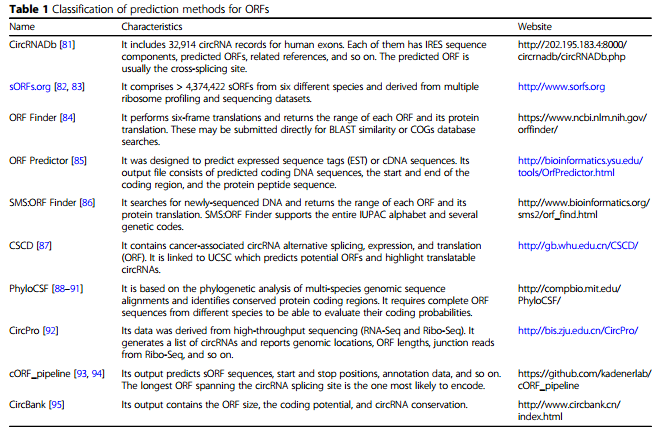
ORF预测:ORF是从ATG(或RNA中的AUG)开始并以三碱基组连续到终止密码子的核酸序列,sORF通常小于300 nt。作者在表1中列举了目前预测ORF的软件和网站;
IRES 预测:IRES是一种RNA调节元件,可募集核糖体,实现核糖体组装和阅读框蛋白质翻译,并启动独立于5’帽结构和直接翻译的蛋白质翻译。IRES介导的翻译的选择性可以调节生理和病理过程,例如细胞生长、增殖、分化、应激反应和凋亡。作者在表2中列出用于预测IRES的网站。

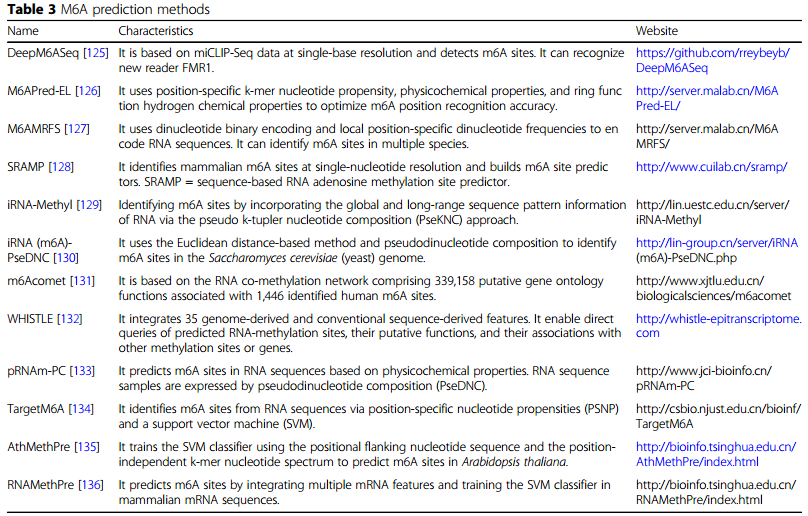
m6A修饰预测:在高等生物的mRNA和ncRNA中,m6A修饰非常普遍。m6A修饰可调节哺乳动物基因的表达,以及转录后水平的RNA稳定性、定位、剪切和翻译。最近发现,m6A对翻译有多种影响,其调节机制异常与肿瘤发生有关。作者在表3中列出筛选m6A基序的常用工具。
保守性分析:保守序列表明潜在的功能和/或在细胞发育和调控中起重要作用。进化保守性(包括ncRNA序列,sORF和小肽氨基酸序列)可作为ncRNA编码功能分析的预测因子。可通过BLAST和UCSC等网站以及MegAlign、MEGA和Clustal之类的软件分析序列的保守性。
另外,目前常见的翻译组学和蛋白质组学也可用于预测ncRNA是否翻译成多肽。
本文也简单介绍了几种实验方法识别ncRNA的翻译功能,例如,将体外构建的FLAG标记的表达载体导入细胞,随后通过蛋白质印迹法在预期的分子量处识别出不同的条带,表明带有FLAG标签的人工构建的ncRNA已被翻译。又如,内源性翻译产物可以通过蛋白质印迹法鉴定,也可以通过特异性抗体来鉴定。分离样品中的蛋白质和多肽,并通过LC-MS / MS测定等。其他方法可以参考原文。
近年来,对编码ncRNA的蛋白质的研究一直在增加,本文也重点介绍了与肿瘤相关的lncRNA和circRNA编码的多肽的功能及作用机制。
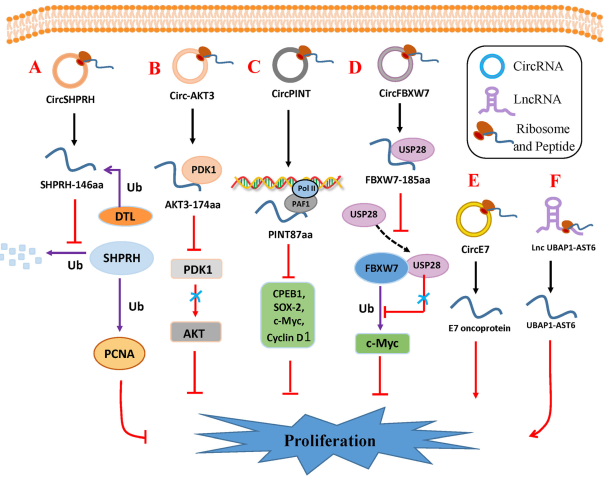
图1 circRNA和lncRNA编码的小肽调节肿瘤增值
1. SHPRH-146aa
SHPRH基因的环状形式编码蛋白质SHPRH-146aa。Circ-SHPRH和SHPRH-146aa在正常人脑组织中高表达,在胶质母细胞瘤中下调。circ-SHPRH中的环化形成串联终止密码子UGAUGA。通过以重叠的遗传密码开始和停止翻译,整个circ-SHPRH被翻译成146-aa蛋白质。针对跨越剪接位点的ORF产生的独特氨基酸序列的抗体,以及通过LC-MS/MS证实circ-SHPRH被翻译为SHPRH-146aa。SHPRH-146aa通过调节蛋白质泛素化途径参与中枢神经系统癌症的发展。 SHPRH-146aa在U251和U373胶质母细胞瘤细胞中的过度表达可降低其体内外的恶性和致瘤性。SHPRH-146aa保护全长SHPRH免受泛素蛋白酶降解。它还通过增殖细胞核抗原来稳定SHPRH,可以抑制细胞增殖和致瘤性(图1a)。
2. AKT3-174aa
Circ-AKT3是由AKT3第三至第七外显子环化形成的。它长524-nt,主要定位于细胞质。由活跃的IRES驱动,circ-AKT3通过重叠的起始-终止密码子UAAUGA编码174-aa蛋白,即AKT3-174aa。AKT3-174aa与AKT3的残基62-232具有相同的氨基酸序列。与正常脑组织相比,胶质母细胞瘤组织中AKT3-174aa被下调。AKT3-174aa,但不是circ-AKT3,起着抑癌作用。 AKT3-174aa过表达可抑制胶质母细胞瘤细胞增殖,辐射抗性和致瘤性。PI3K / Akt途径在各种胶质母细胞瘤发展和进展的致癌信号途径中起着重要作用。PI3K激活后,Akt通过PH域募集到膜上,并在Thr308和Ser473依次磷酸化后被完全激活。PDK1直接使Thr308处的Akt磷酸化。第一步是Akt激活中最关键的步骤。AKT3-174aa的氨基酸序列与AKT3的氨基酸序列部分相同。因此,AKT3-174aa与活化的PDK1竞争性相互作用,抑制Thr308处的Akt磷酸化,并负调控PI3K / Akt信号通路(图1b)。
3. PINT87aa
张等通过对正常人星形胶质细胞和U251胶质母细胞瘤细胞进行circRNA转录组和RNC-RNA测序以及生物信息学整合分析,鉴定lncRNA LINC-PINT的第二个外显子通过自环化形成了环状分子circPINT。后者包含一个sORF和一个天然IRES,其编码从内源性circPINT外显子2而非线性LINCPINT翻译的87-aa多肽(PINT87aa)。它主要位于细胞核,与PAF1直接相互作用,调节PAF1/POLII复合物,抑制下游癌基因cpeb1,sox-2,c-Myc,cyclin D1等的转录延伸,并抑制胶质母细胞瘤和其他癌细胞类型的增殖(图1c)。
4. FBXW7-185aa
Circ-FBXW7的ORF可能跨越剪接位点。它在不同物种之间是高度保守的,并且编码由IRES驱动的185-aa蛋白(FBXW7-185aa),与5’帽的翻译机制无关。使用含有FLAG序列的构建体,可以在ORF终止密码子之前在人细胞中翻译Circ-FBXW7。FBXW7a是FBXW7最丰富的同工型,使用c-Myc作为泛素诱导的降解的肿瘤发生调节剂。泛素化酶USP28通过与FBXW7a的N末端相互作用与c-Myc结合来稳定c-Myc。FBXW7-185aa较FBXW7a而言对USP28的亲和力较高,竞争性地抑制USP28与FBXW7a的结合,因此,干扰USP28诱导的c-Myc稳定,并缩短其半衰期(图1d)。
5. E7 protein
人乳头瘤病毒产生由整个E7 癌基因的ORF产生的circRNA–CircE7。CircE7被m6A修饰,主要定位于细胞质。它与多核糖体紧密相关,可以翻译成E7癌蛋白。在CaSki宫颈癌细胞中敲除CircE7可降低E7蛋白水平,抑制癌细胞增殖和集落形成,并抑制肿瘤生长和恶性肿瘤。 CircE7对于在CaSki子宫颈癌细胞中体外和移植的肿瘤中E7蛋白表达和转化都是必不可少的(图1e)。
6. UBAP1-AST6
蛋白质UBAP1-AST6是从lncRNA翻译而来,主要定位于细胞核,并在A549肺癌细胞中表达。 UBAP1-AST6促进癌症,其过表达显著诱导癌细胞增殖和集落形成(图1f)。
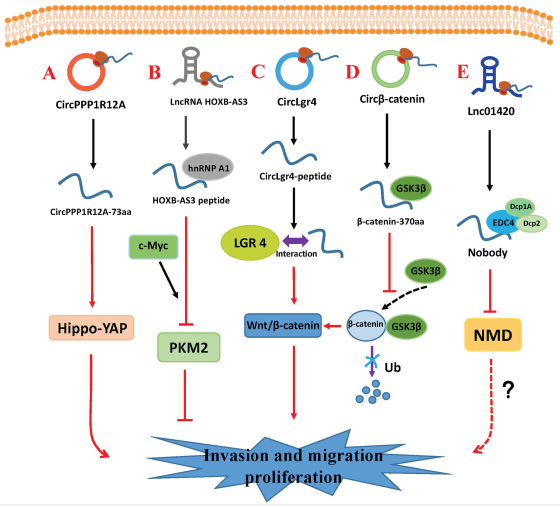
图2 circRNA和lncRNA编码的小肽调节肿瘤的侵袭,转移和增殖
7. circPPP1R12A-73aa
CircPPP1R12A在结肠癌组织中高度表达,可作为生存的预后指标。circPPP1R12A包含一个短的216-nt ORF,编码保守的73-aa肽(circPPP1R12A-73aa)。沉默CircPPP1R12A可显著抑制结肠癌细胞的增殖,迁移和侵袭。具有起始密码子突变ATG/ACG的FLAG-circPPP1R12A过表达载体的构建证实,是circPPP1R12A-73aa而不是circPPP1R12A在结肠癌细胞的增殖,侵袭和转移中起关键作用。通过激活Hippo-YAP信号通路,circPPP1R12A-73aa诱导结肠癌生长和转移(图2a)。
8. HOXB-AS3 peptide
lncRNA HOXB-AS3是一个肿瘤抑制因子,在高度转移性和原发性结直肠癌(CRC)组织中显著下调。HOXB-AS3结合核糖体并编码高度保守的53-aa肽(HOXB-AS3)。HOXB-AS3是内源的,天然存在的,并且在各种肿瘤组织中广泛表达。HOXB-AS3抑制癌细胞的增殖,侵袭和转移并抑制肿瘤的生长。HOXB-AS3水平低的结肠癌患者一般预后较差。HOXB-AS3在hnRNP A1的RGG基序中竞争性结合精氨酸,阻止hnRNP A1与丙酮酸激酶M(PKM)EI9序列结合,拮抗hnRNP A1介导的PKM剪接调控,并抑制PKM 2亚型的形成和miR-18a的产生。HOXB-AS3下调PKM2,但上调PKM1。PKM2是关键调节器有氧糖酵解和增加乳酸生产。因此,HOXB-AS3抑制CRC细胞中的有氧糖酵解(图2b)。
9. CircLgr4-peptide
CircLgr4在晚期CRC中高度表达,并与不良预后相关。 LGR4在大肠肿瘤中也高表达,并通过泛素化和FZD受体稳定作用激活Wnt /β-catenin信号传导。 因此,它驱动大肠干细胞的自我更新和侵袭。 CircLgr4编码circLgr4-肽,该肽与LGR4相互作用以激活LGR4-Wnt信号通路。 CircLgr4以依赖LGR4的方式驱动大肠干细胞的自我更新和侵袭。 circLgr4-肽-Lgr4轴可用于靶向CRC治疗[176](图2c)。
10. β-catenin-370aa
Circβ-catenin衍生自CTTNB1, 具有ORF和IRES,编码β-catenin-370aa,与370-aaβ-catenin互为异构体。具有起始密码子突变的circβ-catenin表达载体的构建表明,其功能可以归因于其蛋白编码能力而不是其非编码特性。Circβ-catenin基因敲低对CTTNB1 mRNA水平无影响,但可显著降低β-catenin蛋白水平。β-catenin被GSK3β磷酸化后,被泛素连接酶β-TrCP泛素化,并被蛋白酶体降解。由circβ-catenin编码的β-catenin-370aa与GSK3β相互作用并起诱饵作用,以阻止其与全长β-catenin蛋白结合,从而抑制了GSK3β诱导的β-catenin降解。在肝癌中,β-catenin-370aa通过减少泛素化,激活Wnt /β-catenin途径并促进肿瘤生长来稳定β-catenin(图2d)。
11. Nobody
LINC01420是在鼻咽癌中高表达的lncRNA。LINC01420表达升高的鼻咽癌患者的总生存率较低。 敲低LINC01420显著抑制鼻咽癌细胞的侵袭。LINC01420 / LOC550643的sORF编码一种序列高度保守的微蛋白,名为nobody。它与mRNA封端蛋白相互作用,直接结合EDC4,从mRNA去除5’帽,促进5’到3’的衰变,并调节正常和异常转录本的降解。nobody主要位于P-body。它的水平随着P-body数目的增加而降低。然而,该过程对肿瘤生长,发育和代谢的影响尚不清楚(图2e)。
结论和未来展望
NcRNA编码的蛋白质引起了很多科学家的好奇心。研究已经确定了存在并证实了ncRNA编码的功能性肽的重要性。但是,对ncRNA编码潜力的评估很困难。用于预测ncRNA中ORF,IRES和m6A的种间保守性的数据库不完整,并且实验验证方法仍在开发中。大多数circRNAs由蛋白质编码的外显子产生,外显子可能与其相关的mRNA重叠,导致难以区分翻译产物的来源。此外,细胞和组织特异性表达使分析变得复杂。因此,可翻译的sORFs的实际数量及其生物学功能仍然未知。
总之,这篇文章总结了ncRNA编码的调节人类癌症行为的小肽的最新进展。这项研究提供了有关ncRNA功能和机制的新观点。因此,还建议未来可能对ncRNA进行更深入的研究,包括是否有更多的功能性肽或ncRNA编码的蛋白,或者早期研究的ncRNA是作为asRNA进行分析还是对其潜在的编码功能进行了研究,其机理是什么?在编码功能肽的ncRNA的动态翻译中,编码小肽的ncRNA是以与RNA相似的方式进行翻译后修饰?哪些因素和条件会影响ncRNA的翻译?
参考文献
Wu, P., Mo, Y., Peng, M., Tang, T., Zhong, Y., Deng, X., et al. 2020. Emerging role of tumor-related functional peptides encoded by lncRNA and circRNA. Molecular Cancer, 19(1), 22.