环状RNA(circRNAs)属于非编码RNA的一个亚类,通过反向剪切形成[1]。其中,通过母基因的5’端及3’端连接在一起形成环状结构。早期的研究认为,circRNA是不可编码的[2],然而,随着测序技术及生物信息学算法的迭代更新,已经发现了许多具有潜在功能的circRNA[3]。在肾细胞癌(RCC)中,hsa_circ_0057105具有双重作用调控机制。一方面,它通过激活EMT上皮间质转化促进RCC细胞的转移。另一方面,它也增加了RCC细胞的铁死亡敏感性。这表明在hsa_circ_0057105的调节下,RCC细胞需要在“攻击性”和“氧化敏感性”之间做出选择。
 一、高通量测序鉴定EMT相关的差异表达circRNA及hsa_circ_0057105表征
一、高通量测序鉴定EMT相关的差异表达circRNA及hsa_circ_0057105表征
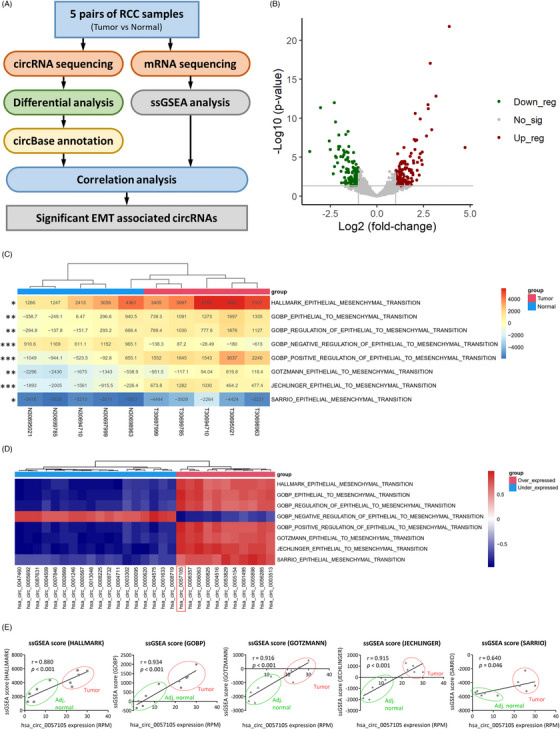 图1. 高通量测序鉴定上皮-间质转化(EMT)相关circRNA
图1. 高通量测序鉴定上皮-间质转化(EMT)相关circRNA
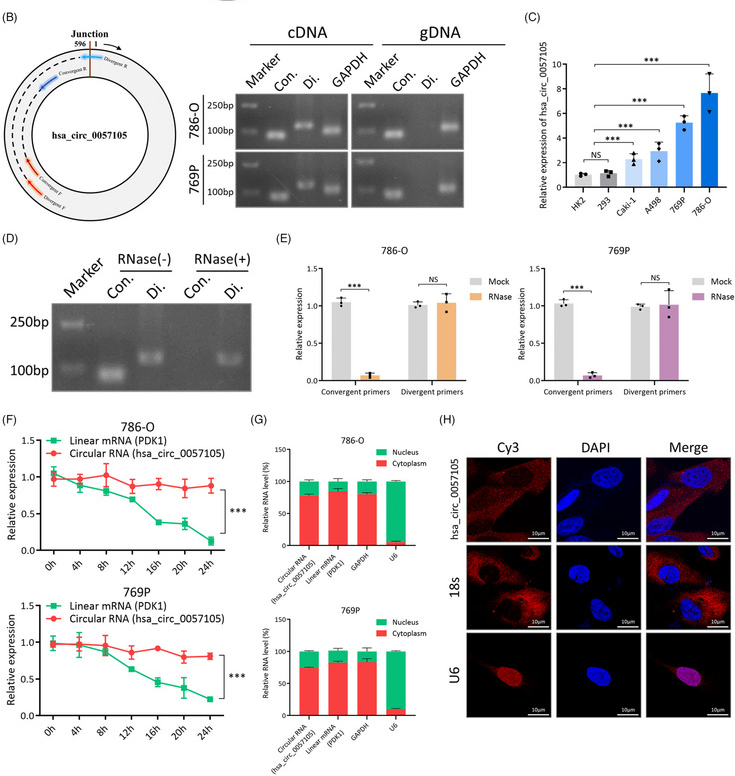 图2. hsa_circ_0057105在肾细胞癌(RCC)中的表达特征
图2. hsa_circ_0057105在肾细胞癌(RCC)中的表达特征
二、多样本下hsa_circ_0057105的表征
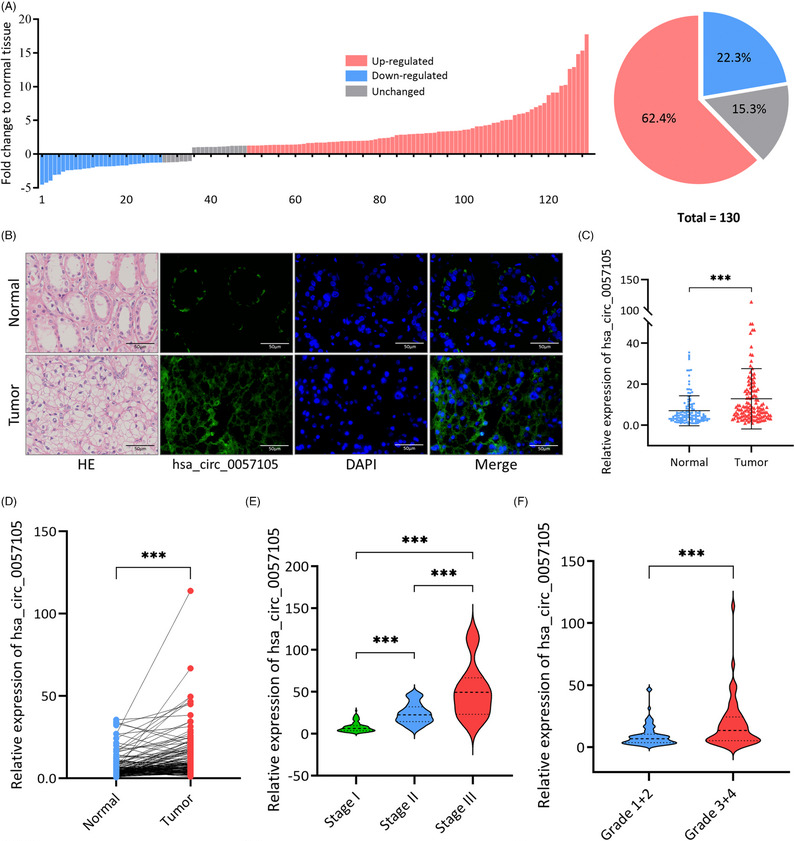 图3. hsa_circ_0057105是肾细胞癌(RCC)中主效致癌因子
图3. hsa_circ_0057105是肾细胞癌(RCC)中主效致癌因子
三、hsa_circ_0057105在RCC细胞系中发挥作用及分子海绵机制
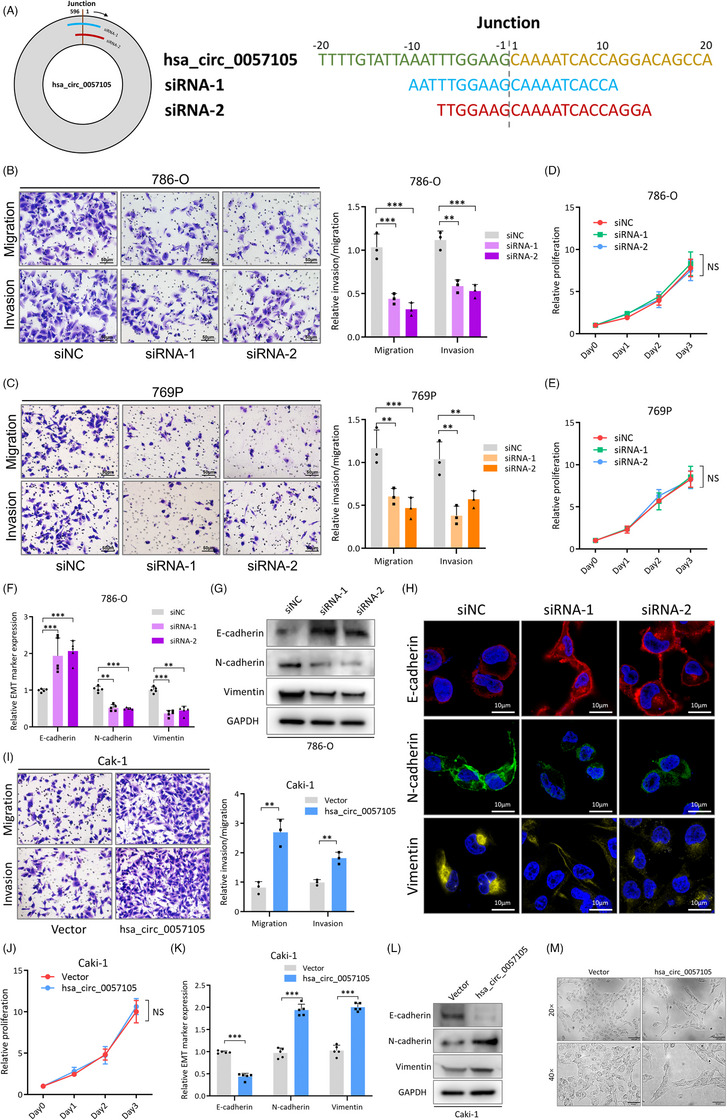 图4. hsa_circ_0057105促进RCC细胞侵袭
图4. hsa_circ_0057105促进RCC细胞侵袭
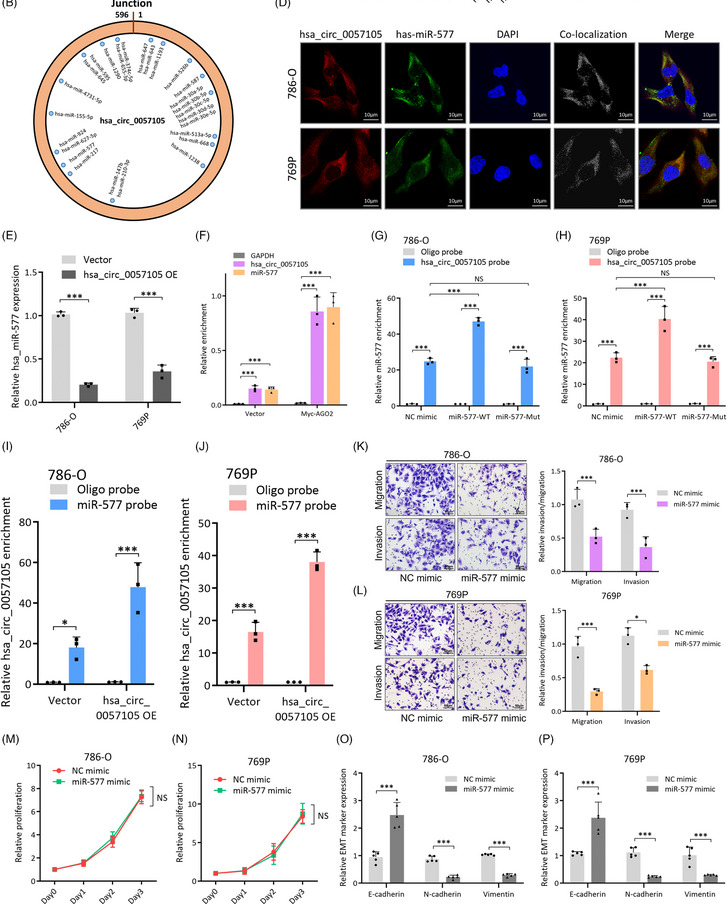 图5. hsa_circ_0057105调控RCC细胞的海绵机制
图5. hsa_circ_0057105调控RCC细胞的海绵机制
四、RCC细胞系中miR-577分子海绵机制下游探索
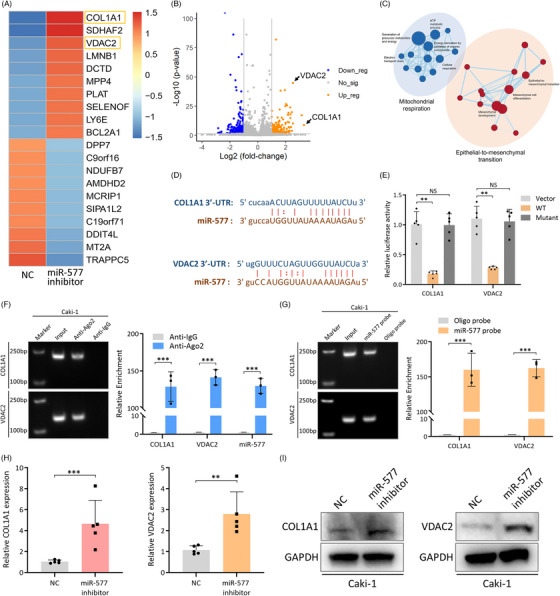 图6. hsa_circ_0057105调控miR-557/COL1A1和VDAC2海绵机制
图6. hsa_circ_0057105调控miR-557/COL1A1和VDAC2海绵机制
五、RCC细胞系中COL1A1及VDAC2靶基因效应
 图7. COL1A1控制RCC细胞上皮-间质转化EMT通路的激活
图7. COL1A1控制RCC细胞上皮-间质转化EMT通路的激活
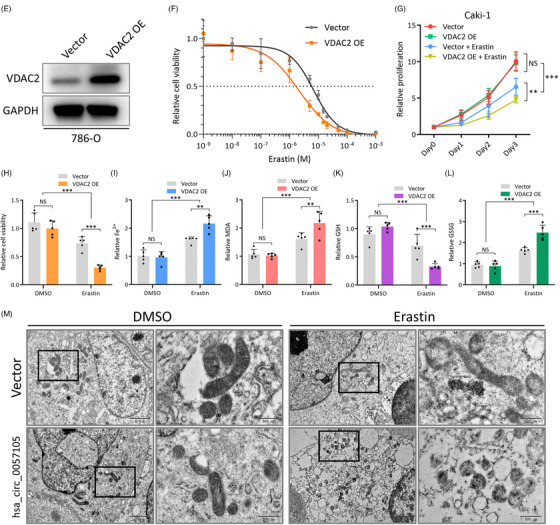 图8. VDAC2促进RCC细胞对铁死亡诱导剂的敏感性
图8. VDAC2促进RCC细胞对铁死亡诱导剂的敏感性
六、hsa_circ_0057105调控miR-577/COL1A1/VDAC2通路来平衡RCC细胞和生物体内的EMT及铁死亡诱导机制
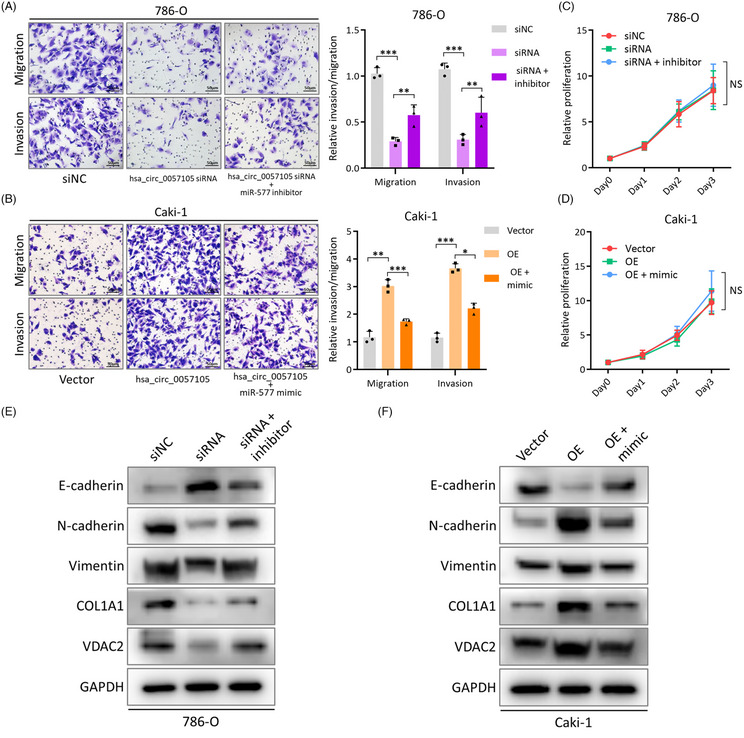 图9. hsa_circ_0057105在RCC细胞中激活EMT并促进铁死亡敏感性
图9. hsa_circ_0057105在RCC细胞中激活EMT并促进铁死亡敏感性
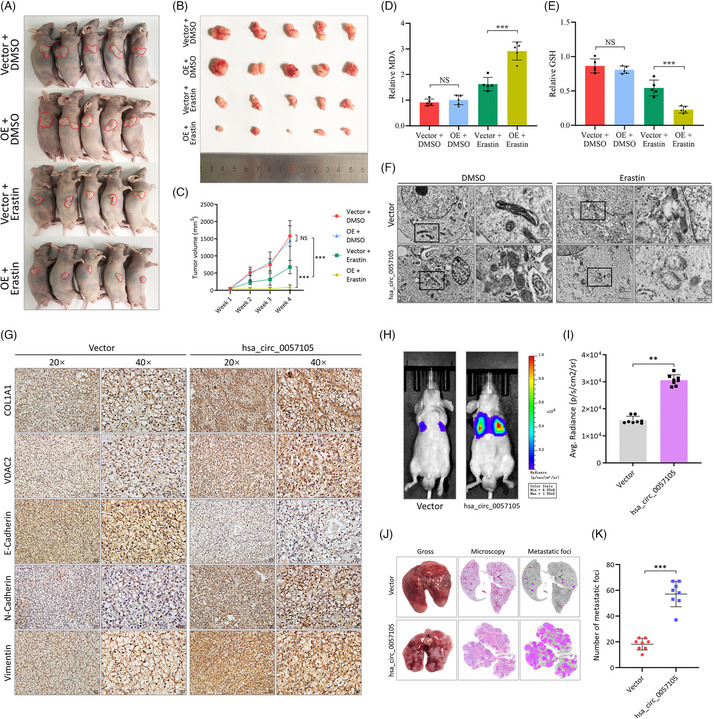 图10. hsa_circ_0057105在小鼠体内激活EMT通路及增强铁死亡诱导剂的敏感性
图10. hsa_circ_0057105在小鼠体内激活EMT通路及增强铁死亡诱导剂的敏感性
总结
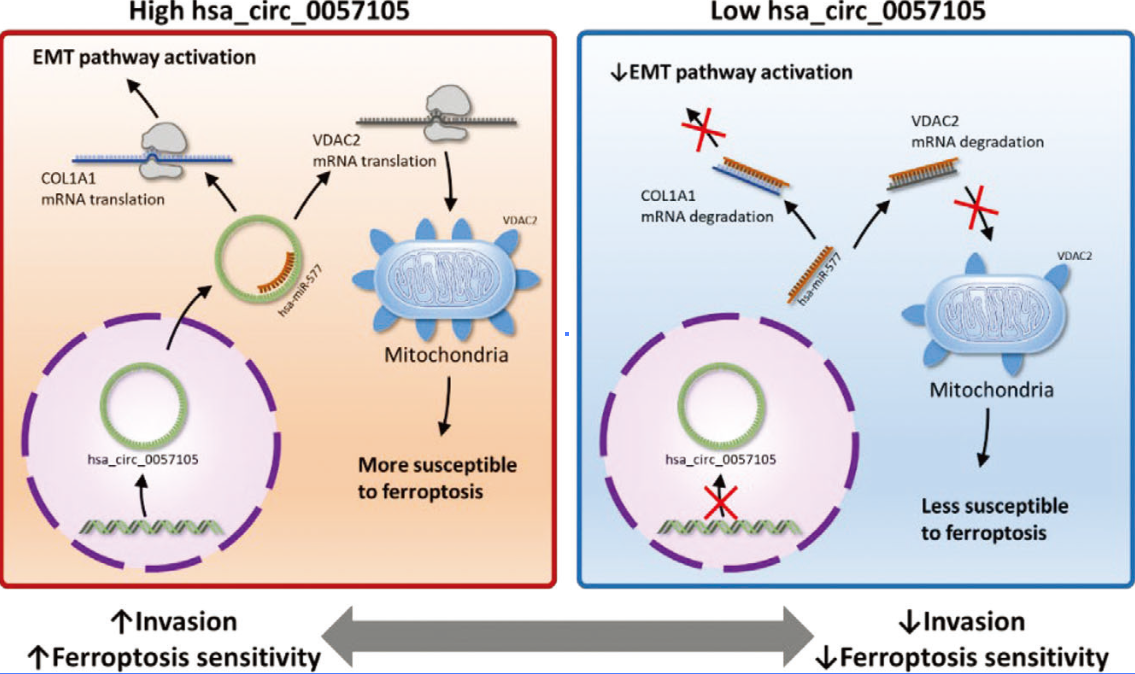 图11. hsa_circ_0057105在RCC中潜在的分子调控机制
图11. hsa_circ_0057105在RCC中潜在的分子调控机制
参考文献:
[2] Cocquerelle C, Mascrez B, Hetuin D, Bailleul B. Mis-splicing yields circular RNA molecules. FASEB J. 1993;7(1):155-160.
[3] Salzman J, Chen RE, Olsen MN, Wang PL, Brown PO. Celltype specific features of circular RNA expression. PLoS Genet.2013;9(9):e1003777.
